|
MAIN PAGE
> Back to contents
Physics of biology and medicine
Reference:
Binhi V., Rubin A.B.
On the quantum nature of magnetic phenomena in biology
// Physics of biology and medicine.
2023. № 1.
P. 44-73.
DOI: 10.7256/2730-0560.2023.1.40435 EDN: SVLAQR URL: https://en.nbpublish.com/library_read_article.php?id=40435
On the quantum nature of magnetic phenomena in biology
Bingi Vladimir
ORCID: 0000-0003-1341-9591
Doctor of Physics and Mathematics
Head of Laboratory, Prokhorov General Physics Institute of the Russian Academy of Sciences
38 Vavilova str., Moscow, 119991, Russia

|
vnbin@mail.ru
|
|
 |
Other publications by this author
|
|
|
Rubin Andrei Borisovich
Doctor of Biology
Head of Department, Lomonosov Moscow State University, Academician of the RAS
119234, Russia, Moscow, Leninskie Gory str., 1/12
|
|
rubin@biophys.msu.ru
|
|
 |
|
DOI: 10.7256/2730-0560.2023.1.40435
EDN: SVLAQR
Received:
12-04-2023
Published:
08-06-2023
Abstract:
The review discusses the microscopic mechanisms of the action of weak magnetic fields on organisms. Magnetobiology distinguishes between magnetoreception, i.e., the effect of a magnetic field on specialized receptors, and a nonspecific response that develops without such receptors. The nonspecific effects of weak magnetic fields are highly general and universal: they occur in all organisms. Often these effects are disguised as the result of the action of uncontrolled random factors, appear as an increased scatter of measurements, and accompanied by low reproducibility. The nature of nonspecific magnetic effects, as is shown in this review, is related to the quantum dynamics of the magnetic moments of electrons, magnetic nuclei, and, possibly, rotations of molecular groups. Among the most substantiated is the spin-chemical mechanism, first of all. Its known low sensitivity to weak magnetic fields can be increased by including spin-correlated radical pairs in the enzymes that catalyze biopolymer processes, e.g., ribosomal ones. We show that research on the effects of significantly weakened magnetic fields compared with the geomagnetic field on cellular processes has prospects for various practical applications. The mechanisms proposed to explain nonspecific effects, but turned out to be untenable, are listed.
Keywords:
quantum effect, magnetobiology, weak magnetic field, hypomagnetic field, nonspecific effect, random effect, molecular mechanism, kT problem, spin chemistry, radical pair mechanism
This article is automatically translated.
You can find original text of the article here.
Introduction The pioneer of the scientific study of the biological effects of the magnetic field (MP) is, apparently, our compatriot Academician A.F. von Middendorf (1815-1894, Russian Empire). Based on his observations of migratory birds, he wrote: "The internal magnetic sense of our 'sky sailboats' takes over the function performed by the compass on ships" [1] p. 9, Fig. 1. 
Fig. 1. The title page of the monograph by A.F. von Middendorf with the materials of the expedition to Siberia. The practical study of the biological effects of MP is initially associated with medicine. It is known, for example, that already in the 1750s German doctors were studying the therapeutic potential of MP [2]. There is evidence in the literature that in the XIX and the first half of the XX centuries, dozens of doctors, including domestic ones, S.P. Botkin, A.V. Seleznev, V.I. Karmilov, etc., tried to use permanent magnets and artificial permanent and variable MP for physiotherapy purposes [3,4]. However, the origin of research into the nature of the phenomenon was associated with the development in the USSR and the USA in the 1960s of emitters of electromagnetic waves in the millimeter range. It turned out that microwaves are able to act on plants and microorganisms [5-7], which is confirmed by numerous modern studies, see e.g. [8-10]. This seemed paradoxical, since the quantum energy of such a field was two orders of magnitude less than the activation energy of chemical transformations, and the frequency selectivity of the action and the low radiation power excluded the thermal nature of the effect. In the development of these studies, already in the 1970s and 1980s, biological effects of weak low-frequency MP were observed, mainly below the level of geomagnetic field intensity (geoMP) and in the frequency range from units to hundreds of Hz [3.11]. The question arose: do the background MP accompanying the transmission and use of electricity affect health? In 1983 , in order to coordinate research on the nature of this phenomenon and the development of therapeutic methods , the The Problem Commission of the Ministry of Health of the USSR "Magnetobiology and magnetotherapy in medicine" [12]. In 1992, the US Congress approved a five—year research program — the study of electric and magnetic fields and the dissemination of public information - worth more than $ 40 million [13]. It should be noted that by the time the program was completed in 1998, the issue of the impact of MP on human health had not been resolved unequivocally. Only the subsequent decades of collective efforts of researchers around the world allowed us to establish the approximate degree of danger of background MP [14] p. 332, [15-20]. Currently, it has been reliably established that MP orders of magnitude smaller than geoMP can cause biological effects [21-26]. More than ten thousand scientific papers have been published on the effect of weak MP on organisms. Nevertheless, such an action continues to raise questions. Today, magnetobiology distinguishes between specific — due to special receptors — and non-specific magnetic effects [27,28]. Specific effects occur due to specialized magnetic receptors developed during biological evolution. They ensure the survival of some animal species during long seasonal migrations. There are dozens of experiments on animal navigation, where in large enclosures or swimming pools, magnetic conditions characteristic of places of seasonal stays separated by several hundred kilometers were simulated. At the same time, the animals radically changed the direction of movement [29]. In other observations, magnetic storms changed the direction of migrations [30], etc. The facts of sensitivity of some organisms to MP variations of the order of geomagnetic variations are reliably established. A specific magnetic effect is a magnetic sense, i.e. magnetoreception [21,31]. For example, some migratory birds have a special receptor combined with the visual system, Fig. 2. A bird can "see" changes in MP of the order of tens of NT and use this for orientation and navigation along the magnetic relief of the Earth [32]. Some migrating insects have a similar ability, also related to vision [33]. 
Fig. 2. The robin Erithacus rubecula, which is believed to use a quantum magnetochemical compass for orientation and navigation in geoMP during seasonal flights at distances up to 2000 km. Photo © F.C. Franklin / CC-BY-SA-3.0. In addition to the effects of magnetic navigation, variations of MP of the order of 1 µT have an effect on organisms that do not have special receptors. These are the nonspecific magnetic effects that are causing more and more interest. Non-specific effects are not magnetoreception, since such presupposes the presence of receptors. Even 30 years ago, MP was considered a factor acting on a person bypassing the senses [4], that is, bypassing specialized receptors. Nonspecific effects are observed in many organisms, from protozoa and fungi to insects, plants [34,35], Fig. 3, fish, animals and humans [36]. Interest in these effects is increasing, as MP can change many properties. In particular, gene expression changes [37,38]. In other words, MP may be one of the factors controlling protein synthesis. However, it is not yet possible to use the full power of this method of gene control, since the nature of the primary biophysical sensor MP, which is not a magnetoreceptor, has not yet been explained in the body. No such sensor has been detected experimentally. 
Fig. 3. Arabidopsis thaliana watercress does not have special magnetic receptors, but the properties of its seedlings change in weak MP. The photo was kindly provided by Prof. A. Batschauer (Alfred Batschauer). From 1980 to the present, many physical mechanisms and mathematical models have been proposed to explain the biological effect of weak MP. The list could include those that take into account: biogenic magnetite and contamination by magnetic nanoparticles; thermal heating and eddy currents; magnetohydrodynamic effects and chemical kinetics with bifurcations; cyclotron, parametric and stochastic resonances; phase transitions in magnetic fields and spin chemistry effects; interference of quantum states of ions and molecular groups; coherent excitations; metastable states of water, etc. [22]. To date, only a few approaches have been preserved and developed, but they also remain hypotheses. In recent decades, several reviews of theoretical works in the field of magnetoreception have been published [32,39]. However, there have been no reviews of theoretical concepts aimed at explaining non-specific effects. To fill this gap is the purpose of this work.
The following section discusses the problems that arise in explaining the biological effects of weak MP. It is followed by a section in which it is proved that plausible mechanisms of the magnetic response of organisms take into account the quantum dynamics of the primary acceptors of MP — magnetic moments of electrons, first of all. The possible quantum mechanisms are listed and the essence of the spin-chemical mechanism of radical pairs (RPM), the mechanism most represented in the scientific literature, is explained. Special sections are devoted to a more detailed discussion of the difficulties of this mechanism, due to the short time of thermal spin relaxation, and discussion of the mechanism of statistical amplification of weak primary MP signals in the process of cellular translation, a mechanism that may relieve the difficulties of RPM. This is followed by a section in which it is shown that the experimental determination of the mechanism of biological action of a weak MP is most appropriate to be carried out using a hypomagnetic field. The results of the review are concentrated in the Conclusion. Appendices 1 and 2 introduce quantum mechanisms alternative to RPMs; Appendix 3 contains descriptions of frequently proposed but untenable mechanisms of biological action of weak MP; it is explained why they are untenable. The review is addressed to the reader who is not yet familiar with this area of research. Accordingly, the style of presentation of the material is simplified. This makes it possible to illustrate complex quantum phenomena in terms of classical physics. At the same time, the inevitably limited scope of the review can only give a general idea of magnetic biological phenomena. A deeper understanding of them will require referring to specialized sources from the list of references. Problems of explaining the biological effects of weak MP Just as an electric field interacts with electric charges, an MP interacts with magnetic moments. Any molecular mechanism of magnetic effects in organisms is based on this. The magnetic moment is a physical quantity describing the magnetic properties of an object. It is known, for example, that an electron has both a mechanical moment, or spin, and a magnetic, collinear spin. The classic analogy is a spinning top carrying charges on its rim. The circular motion of the charges creates an MP directed along the axis of the top — this is, in fact, the MP of a microscopic magnet. Its magnitude and direction are set by the magnetic moment vector 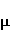 . . In the external MP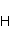 , the magnetic moment behaves like a compass needle. The external MP tries to deploy the moment so that their directions coincide, i.e. so that the energy of their interaction , the magnetic moment behaves like a compass needle. The external MP tries to deploy the moment so that their directions coincide, i.e. so that the energy of their interaction 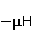 (the GHS system) is minimal. However, this is hindered by the law of conservation of the mechanical angular momentum, which is rigidly connected with the magnetic one. As a result, there is a so-called precession of the magnetic moment in the MP, similar to the precession of the top in the gravity field. This ultimately causes the biological response of the organism to MP. In terms of quantum mechanics, MP, acting on the quantum state of the magnetic moments of electrons, changes their spin state, which, as is known, is a fundamental chemical factor. We should immediately note that processes involving spin are quantum processes that cannot be described in the language of classical physics. (the GHS system) is minimal. However, this is hindered by the law of conservation of the mechanical angular momentum, which is rigidly connected with the magnetic one. As a result, there is a so-called precession of the magnetic moment in the MP, similar to the precession of the top in the gravity field. This ultimately causes the biological response of the organism to MP. In terms of quantum mechanics, MP, acting on the quantum state of the magnetic moments of electrons, changes their spin state, which, as is known, is a fundamental chemical factor. We should immediately note that processes involving spin are quantum processes that cannot be described in the language of classical physics. In biological tissue, magnetic moments have a different nature. In addition to the magnetic spin moments of electrons, these are the magnetic moments of protons and other magnetic nuclei and the moments of the orbital motion of electrons or charged molecular groups. The energy of magnetic moments — the primary "targets" of MP, initiating a biological response — is extremely small, which generates the following discrepancy. The energy, for example, of the electron moment in the geoMP is about 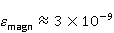 eV. This is seven orders of magnitude lower than the height of the chemical activation barrier eV. This is seven orders of magnitude lower than the height of the chemical activation barrier 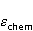 in the range from units to tens in the range from units to tens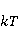 , where , where 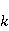 is the Boltzmann constant and is the Boltzmann constant and 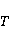 is the temperature of the medium. At the physiological temperature is the temperature of the medium. At the physiological temperature 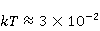 of eV. Physically, is, in order of magnitude, the average thermal energy of eV. Physically, is, in order of magnitude, the average thermal energy 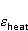 per degree of freedom in a state of thermal equilibrium. Combining the physical and chemical aspects, you can write per degree of freedom in a state of thermal equilibrium. Combining the physical and chemical aspects, you can write 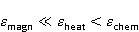 , or , or 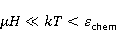
where is the temperature of the nearest environment of the magnetic moment (or is the equilibrium thermal energy of the particle carrying the magnetic moment). The first inequality fixes the disparity in magnitude of the magnetic and thermal forces acting on the particle. These inequalities gave rise to the so-called problem, which for a long time raised doubts about the existence of magnetobiological effects. Indeed, it seems surprising that MP with such a large discrepancy can cause chemical changes.
How is the problem solved ? Looking ahead, we note with respect to the first inequality 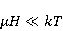 that the magnitude of microscopic magnetic moments that the magnitude of microscopic magnetic moments 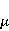 is equal in order of magnitude to the electron magnetic moment or three orders of magnitude less than the nuclear moments. The MP value is equal in order of magnitude to the electron magnetic moment or three orders of magnitude less than the nuclear moments. The MP value 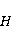 is limited by the geoMP level fields. Therefore, their product is limited by the geoMP level fields. Therefore, their product 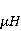 cannot be arbitrarily large. However, to overcome the inequality, the effective temperature of the magnetic moment may be small if the moment is sufficiently well isolated from the disturbing action of the thermostat. However, at the same time, the second inequality takes on the appearance cannot be arbitrarily large. However, to overcome the inequality, the effective temperature of the magnetic moment may be small if the moment is sufficiently well isolated from the disturbing action of the thermostat. However, at the same time, the second inequality takes on the appearance 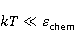 that it seems an insurmountable obstacle to the occurrence of magnetic effects. But this is not the case. There is a class of chemical reactions studied by spin chemistry — reactions involving radical pairs [40] — in which MP is not an energetic, but a controlling factor. The mechanism of such reactions is discussed in the following sections. In short, spin selection rules may prohibit reactions initiated by thermal perturbations of the medium, but the prohibition can be partially lifted by the action of MP on spin magnetic moments. Therefore, it is the first inequality that is the main problem. However, as mentioned above, magnetic moments that are weakly associated with the thermostat remain in conditions of low effective temperature for a relatively long time, — when that it seems an insurmountable obstacle to the occurrence of magnetic effects. But this is not the case. There is a class of chemical reactions studied by spin chemistry — reactions involving radical pairs [40] — in which MP is not an energetic, but a controlling factor. The mechanism of such reactions is discussed in the following sections. In short, spin selection rules may prohibit reactions initiated by thermal perturbations of the medium, but the prohibition can be partially lifted by the action of MP on spin magnetic moments. Therefore, it is the first inequality that is the main problem. However, as mentioned above, magnetic moments that are weakly associated with the thermostat remain in conditions of low effective temperature for a relatively long time, — when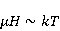 , — and the magnetic effect has time to develop before thermal equilibrium is established as a result of relaxation. , — and the magnetic effect has time to develop before thermal equilibrium is established as a result of relaxation. The above inequalities allow us to eliminate many hypotheses about the nature of magnetic biological effects. These are hypotheses that cannot explain why a weak MP, which changes the energy of magnetic moments by only one ten-millionth part, can cause effects that overcome thermal disturbances of these magnetic moments. In particular, it is sometimes believed that MP, acting on the magnetic moment of a microparticle localized in some oscillatory potential, could lead to an increase in the energy of oscillations to the level and initiate some chemical reaction. Such a mechanism circumvents the problem by accumulating magnetic energy in mechanical vibrations. But its implementation is impossible due to the inertia of mechanical movements. Indeed, in inertial processes, the forced change in the energy of mechanical motion or the coordinate of a particle is proportional to the square of time . Then the small magnetic forces usual for magnetobiology can give energy to a microparticle only for an absurdly long time — even in the most favorable idealized situation and in the absence of thermal disturbances. Since the mechanisms of magnetic effects, formulated in the language of classical physics, necessarily include mechanical motion, modified by MP, they are unreal due to inertia. Although they could occur in strong MP of the order of 1 Tl or more [41], they cannot underlie the nonspecific effects of weak MP. Unfortunately, attempts to explain magnetic effects in biology within the framework of classical physics, other than through the action of MP on the quantum dynamics of magnetic moments, are still being published today. It is important, therefore, to emphasize the failure of such attempts. . Then the small magnetic forces usual for magnetobiology can give energy to a microparticle only for an absurdly long time — even in the most favorable idealized situation and in the absence of thermal disturbances. Since the mechanisms of magnetic effects, formulated in the language of classical physics, necessarily include mechanical motion, modified by MP, they are unreal due to inertia. Although they could occur in strong MP of the order of 1 Tl or more [41], they cannot underlie the nonspecific effects of weak MP. Unfortunately, attempts to explain magnetic effects in biology within the framework of classical physics, other than through the action of MP on the quantum dynamics of magnetic moments, are still being published today. It is important, therefore, to emphasize the failure of such attempts. Since the appearance of the famous book "What is Life?" [42] by one of the founders of quantum mechanics, E. Schrodinger, more and more biological processes have been discovered, the study of which requires a "quantum microscope". In addition to the processes governing mutations and heredity, these are photosynthesis, respiration, vision, smell, enzyme reactions in general and other forms of energy and charge transfer. Even to understand such a complex phenomenon as consciousness, the ideas of quantum physics are attracted today, see the review in [43]. As biophysics and molecular biology master nano and sub-nano ranges of sizes and times, the use of quantum physics becomes inevitable [44]. A corresponding concept, quantum biology, has also been formed, combining studies of biological processes, the description of which in the language of classical physics is not productive [45,46]. Magnetobiological effects are also quantum. The response of organisms to a weak magnetic field cannot be interpreted in any other way than the influence of MP on the dynamics of magnetic moments, the main representative of which in the body is the quantum dynamics of electrons. The next important problem is that non-specific effects are difficult to reproduce. Unlike specific, non-specific effects obtained in one laboratory are difficult to reproduce in another, for which magnetobiology is often criticized. The decrease in reproducibility is explained by the influence of various random factors affecting the outcome of the experiment [47]. Firstly, the values of controlled factors are never strictly constant. Fluctuations of MP, for example, in rooms reach hundreds of nTl and even more near power lines and electric transport [48]. The heterogeneity of MP in biological thermostats reaches tens of µT, which is often overlooked in magnetobiology studies [49], although it is known that some animals are sensitive to MP of 15-30 NT [39,50,51] and even 5 NT [52]. The biological effect of magnetic storms is confirmed by their reproduction in artificial conditions [53,54] or by their compensation [55]. However, in biological laboratories, the level of geomagnetic variations is almost never monitored. Hence the randomness of the measurement results, and hence the reduced reproducibility. There is also a methodological reason for the reduced reproducibility — insufficient adequacy of the statistical methods used to the features of magnetic biological effects [56,57]. Secondly, non-specific effects in magnetobiology depend on factors of different nature [58-60], not all of which can be controlled. Only physical factors are more than ten [56]. Many biological processes are sensitive to changes, for example, temperatures at 0.1oC, although such fluctuations are typical for thermostats. In addition to temperature, MP and electric field, these are humidity, pressure, lighting, the rate of their changes, etc. There are also chemical, physiological and genotypic factors. Finally, the low reproducibility of non-specific effects is caused by randomness that occurs already at the micro level. This is due to the fact that the conversion of the MP signal into a biochemical signal depends on random internal conditions.
Randomness — against the background of homeostasis — manifests itself at all time scales and levels of organization of living matter, starting from the molecular and ending with the level of slow evolutionary processes [61] pp. 281-283. Thermal, chemical and biochemical fluctuations make the internal conditions of the functioning of many biophysical structures, including biophysical sensors of MP, random. Internal conditions are the state of the local environment of MP sensors. These are fluctuating concentrations, activities and steric limitations of protein agents, features of interactions with the thermostat, as well as molecular rotations [62]. Such conditions, for example, for a spin-correlated pair of radicals as a magnetic sensor — see below — are the viscosity of the medium, the details of interactions that determine the time of thermal relaxation of spins, the rate of chemical reaction of radicals, concentrations, etc. The internal conditions in the body are constantly changing due to regulatory processes controlled by a variety of receptor systems and feedbacks. Measurements of biological characteristics fall into different phases of regulatory processes. This means that the response to a change in MP is random due to fluctuations in internal conditions caused by both external and internal regulatory factors. Fluctuations in conditions constantly "turn on" and "turn off" various groups of magnetic biophysical sensors. Consequently, the measurement results obtained with different groups of sensors turned on form not even just a random variable, but a set of realizations of different random variables, i.e. a heterogeneous random effect. We should add that, unlike specialized receptors localized in certain parts of the body, the magnetic moments of electrons and nuclei, spin and orbital, are present in almost all biological molecules distributed throughout the body. Is it possible to say in advance which of them will transmit the MP signal to the level of biochemical reactions and where it will happen? Not yet. In these conditions — with heterogeneity of responses — the concepts of threshold, dose, and MP-dependence in general, have only a limited meaning, which indicates the need for special methods for studying non-specific effects [56]. Finally, we note another problem of the magnetobiology of nonspecific effects — the problem of biological detection of small variations of MP against the background of much larger random fluctuations of the magnetic field in urban conditions. Without solving this problem, it is impossible to explain, for example, the effect of geomagnetic storms, with an intensity rarely exceeding 500 NT, on the functioning of organisms. Biological effects of magnetic storms have been recorded both in the human body and in other organisms [63,64]. Low-frequency fluctuations of GEOMPS, as well as urban magnetic noise, apparently have a direct effect on organisms [30,51,53,65]. Most likely, these facts will not require the involvement of any additional explanatory mechanisms. The explanation is possible on a single basis of the influence of MP on magnetic moments. It is only necessary to take into account the frequency and time factors of magnetic exposure. So, on the one hand, it was noted in [36] that the frequency spectra of geo- and urban magnetic fluctuations are shifted relative to each other by two or three orders of magnitude. It is assumed that the process of forming a biological response to the primary signals of geomagnetic disturbances in the body involves their integration into a system of biochemical signaling devices and regulators. The concentration of molecules whose biosynthesis and decay is slow may reflect the accumulated and averaged effect of weak magnetic signals. On the other hand, it was shown in [48] that the intensity of geomagnetic noise in the low-frequency region can exceed the intensity of urban magnetic noise at night, when mainly the adverse effect of magnetic storms on the human condition is observed. Which mechanisms are most plausible? The low effective temperature, or metastability, of the primary target and the inertia-free response to MP as a condition for the operation of the molecular mechanisms of the action of weak MP on biological systems are essential constraints. Plausible mechanisms should be based, firstly, on the dynamics of magnetic moments. Indeed, the final angular velocity of the precession of magnetic moments is proportional to the applied MP and does not depend on time explicitly. The precession of the moment occurs in the MP immediately and does not require the accumulation of energy of the microparticle. This is a consequence of the independence of the spin energy or mechanical rotation associated with the magnetic moment from the direction of the moment. The direction of angular momentum changes here proportionally, i.e. inertially [47]. Secondly, the dynamics of magnetic moments must be quantum, since it is impossible to get rid of the involvement of spins in magnetic effects. On the other hand, if we talk about angular moments of orbital motion, they are very small, of the same order as the electron spin, and their quantum nature follows from the Heisenberg uncertainty relation. Thirdly, the precession of moments should occur under conditions of slow thermal relaxation, when the effect of thermal disturbances can be neglected. Therefore, the mechanisms of the influence of MP on abstract single magnetic moments [66], on the spin magnetic moments of electrons of radical pairs [28,32,67], on the magnetic moments of nuclei [68,69] and the orbital motion of molecular groups inside proteins and in other biophysical structures [27,70] are plausible. The slowness of thermal relaxation, or a relatively large time interval of coherence of the motion of moments, in these mechanisms has not yet been proven, but is only assumed. More about this will be discussed below. The assumption is gradually developing that proton exchange and metastable states in liquid water could be responsible for non-specific magnetic effects [71-73]. However, these hypothetical mechanisms have not yet acquired the form of mathematical models with predictive power. So far, they do not satisfy one of the principles of scientific methodology — the principle of verifiability. Therefore, we will confine ourselves to discussing the advantages and disadvantages of the above-mentioned mechanisms of MP action on magnetic moments in biophysical structures. These mechanisms arise naturally when considering the general properties of a chemical process. The nonspecific effect of weak MP is, by definition, the effect of MP on the chemical process in the absence of receptors, i.e. under fairly general conditions. There are two main stages in the chemical process: 1) convergence of reagents, often diffusive, at a distance of the order of chemical bond lengths and 2) the actual act of chemical reaction, i.e. rearrangement of electronic shells. At physiological temperature, the energy imparted by magnetic forces is many orders of magnitude less than the energy of diffusion motion. For this reason, the MP cannot influence the course of the process at the first stage. However, MP is able to influence the quantum process of the second stage — the act of reaction. A priori, the influence could be carried out in different ways. The effect could occur through the spin effect on the probability of electron rearrangement. The other way is carried out through one of the steric factors, through the influence of MP on the spatial structure of the electronic wave functions of interacting molecules. Thus, if the key factor is the rearrangement of electronic shells under the action of MP, then the following scenarios are plausible.
 MP affects the rearrangement of electrons, for example, in the reaction of the decomposition of a molecule MP affects the rearrangement of electrons, for example, in the reaction of the decomposition of a molecule 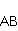 into free radicals into free radicals 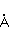 and and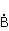 , if the reaction proceeds with the formation of an intermediate , if the reaction proceeds with the formation of an intermediate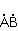
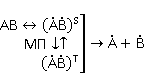
which can be in different spin states. This is a simple example of reactions involving radical couples. Spin chemistry studies such reactions. MP affects the precession (in the vector model) of single moments, for example, spin magnetic moments of electrons. Precession becomes uneven in the variable MP or slows down in the hypomagnetic MP [66], which may affect the probability of acts of adjacent reactions, i.e. also affect the rearrangement of electrons. MP changes the spatial structure or phases of angular quantum states of rotating molecular groups and electron clouds, which leads to interference effects [74]. Unlike the first scenario, or the spin-chemical mechanism of radical pairs (RPM), the last two — the mechanism of mixing quantum levels [66] and the mechanism of the molecular gyroscope [74] — do not have reliable experimental confirmation today. Although they predict a number of observed features of non-specific effects that the RPM cannot explain, they nevertheless remain speculative constructions. The initial information about them is given in Appendices 1 and 2, and below we continue to consider the RPM — the most plausible scenario of nonspecific magnetic effects. Radical pairs occur, for example, in photoliases and cryptochromes under the influence of optical blue and UV radiation. They also occur in many enzyme-substrate reactions [75,76], for example, biopolymer reactions [40], since the redox process often takes place in the active center of the enzyme, accompanied by electron transfer. All this creates a promising basis for explaining magnetic effects in organisms. Let us explain the processes depicted in the above diagram of the occurrence of free radicals. Let a part 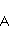 of a complex molecule contain a covalent bond that is broken as a result of electron transfer to a part of a complex molecule contain a covalent bond that is broken as a result of electron transfer to a part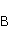 . In the simplest case, the covalent bond connecting and is broken. Immediately after the electron transfer, two radicals are formed, and since the parts of the molecule now contain unpaired electrons. The spin state of the radical pair at the initial moment of time is correlated: the spins of electrons localized on and evolve in a consistent or coherent manner, since random thermal disturbances have not yet had time to destroy coherence. . In the simplest case, the covalent bond connecting and is broken. Immediately after the electron transfer, two radicals are formed, and since the parts of the molecule now contain unpaired electrons. The spin state of the radical pair at the initial moment of time is correlated: the spins of electrons localized on and evolve in a consistent or coherent manner, since random thermal disturbances have not yet had time to destroy coherence. To describe spin states, the so-called Zeeman basis is usually used, 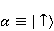 and and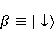 , denoting states with a projection of spin on the MP axis , denoting states with a projection of spin on the MP axis 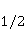 and and 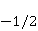 , respectively. One of the basic states of a system of two electrons is then denoted as , respectively. One of the basic states of a system of two electrons is then denoted as 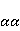 , where the spin state of the first electron is indicated in the first place, and the second is indicated in the second place. Other basic states of the pair are, obviously, , where the spin state of the first electron is indicated in the first place, and the second is indicated in the second place. Other basic states of the pair are, obviously, 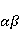 , , 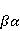 and and 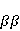 . . The stationary states of the pair are the eigenstates of the energy operator — the Hamiltonian of the system. Stationary states have a certain symmetry in the permutation of particles. If there is only the energy of magnetic moments in MP, then this is an antisymmetric singlet state 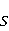 in which the total spin of the pair is zero, in which the total spin of the pair is zero, 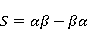 , and three triplet states corresponding to the total spin of one with different spin projections on the direction of MP. Triplet states are symmetric by permutation of a pair of particles: , and three triplet states corresponding to the total spin of one with different spin projections on the direction of MP. Triplet states are symmetric by permutation of a pair of particles: 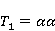 , , 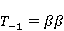 and and 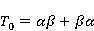 . . The transfer of an electron from one part of a molecule to another, or from one molecular group to another, as in proteins, means the appearance of two paramagnetic electrons in a singlet joint state. This is a consequence of the law of conservation of angular momentum of a system of two electrons (recall that spin is the intrinsic angular mechanical moment of a particle). The total angular momentum of a pair of electrons as a result of the transfer usually does not change, and before the transfer it was zero, since it belonged to paired electrons of a covalent bond.
In some cases, the intermediate resulting from electron transfer and initially being in the singlet state 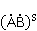 lives long enough, and the MP can affect the evolution of its spin state, or, what is the same, change the speed of singlet-triplet (CT) transitions lives long enough, and the MP can affect the evolution of its spin state, or, what is the same, change the speed of singlet-triplet (CT) transitions 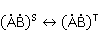 occurring for various reasons. Since the recombination of parts of the intermediate into the initial form is possible only from the singlet state, CT transitions, first reducing the proportion of the singlet state, reduce the probability of recombination and, therefore, increase the probability of the formation of free radicals and . This was the basis for frequent assumptions that MP increases the concentration of free radicals in the body and leads to various negative effects. However, the processes of transfer with the appearance of radical pairs in the body are more complicated than the above illustrative scheme, and the possible relative increase in the concentration of free radicals, if it occurs, is very small and does not explain the observed effect of weak MP on organisms. This will be explained below. occurring for various reasons. Since the recombination of parts of the intermediate into the initial form is possible only from the singlet state, CT transitions, first reducing the proportion of the singlet state, reduce the probability of recombination and, therefore, increase the probability of the formation of free radicals and . This was the basis for frequent assumptions that MP increases the concentration of free radicals in the body and leads to various negative effects. However, the processes of transfer with the appearance of radical pairs in the body are more complicated than the above illustrative scheme, and the possible relative increase in the concentration of free radicals, if it occurs, is very small and does not explain the observed effect of weak MP on organisms. This will be explained below. In assessing the plausibility of the RPM, it is important to take into account thermal disturbances that destroy the consistency, or coherence, of the dynamics of magnetic moments or electron spins. ST states of electron spins make sense (description in terms of ST states is effective) only if the dynamics of the spins is coherent. Coherence is destroyed by thermal fluctuations. In the thermalized state, the motion of the spins is essentially chaotic. MP is not able to influence the evolution of a pair of spins, or the CT conversion. Magnetic effects are possible only at the interval of spin coherence, or during  the relaxation time of spin coherence, Fig. 4. the relaxation time of spin coherence, Fig. 4. 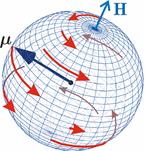
Fig. 4. The nature of the precession of the "vector" of the spin magnetic moment 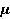 in the MP in the MP 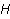 under the action of thermal disturbances. The red arrows represent the trajectories of the end of the vector in the intervals between heat strokes. Precession is coherent in these areas. At the end of each such section, a fast random jump of the vector to the beginning of some other section occurs. If thermal disturbances are small, then impacts are rare, and precession is mostly coherent. If the perturbations are large, then the coherence areas are very small and the motion is indistinguishable from random. In this state, the MP does not affect the evolution of the magnetic moment. It should be borne in mind that the vector spin model is only a convenient illustration of the quantum mechanical process, since there is no spin vector as such. under the action of thermal disturbances. The red arrows represent the trajectories of the end of the vector in the intervals between heat strokes. Precession is coherent in these areas. At the end of each such section, a fast random jump of the vector to the beginning of some other section occurs. If thermal disturbances are small, then impacts are rare, and precession is mostly coherent. If the perturbations are large, then the coherence areas are very small and the motion is indistinguishable from random. In this state, the MP does not affect the evolution of the magnetic moment. It should be borne in mind that the vector spin model is only a convenient illustration of the quantum mechanical process, since there is no spin vector as such. In the theory of spin relaxation, the longitudinal and transverse relaxation times are distinguished. We are interested in orders of magnitude, so we use the generalizing concept of the characteristic time of thermal spin relaxation. In most RMP models of magnetic navigation of animals, the spin relaxation time is assumed to be very large, — more than all other characteristic time scales of relevant processes. Therefore, the effects of thermalization are simply not taken into account. Since the grounds for such idealization are highly controversial, we further discuss the contribution of spin thermalization in more detail. But first we will make one significant clarification regarding the description of quantum systems in terms of CT states. In the experiment, an increase in the concentration of reactive oxygen species, in particular singlet oxygen, was observed under the action of relatively weak MP. Unlike the triplet ground state of molecular oxygen, its singlet form with the lowest energy, called singlet oxygen, has a high reactivity and can damage biopolymers [77]. For this reason, it is sometimes assumed that MP affects the body by directly stimulating CT transitions and increasing the concentration of singlet oxygen. This assumption is incorrect. The energy difference between the C- and T-states of the oxygen molecule corresponds to the energy of the near-IR radiation quanta of about 1280 nm, which is too much for the effects of weak MP to occur. Let's consider this circumstance in more detail. The CT states of molecular terms differ significantly from the CT states of a pair of radicals, which is a consequence of the huge difference in the magnitude of the exchange interaction of electrons in an oxygen molecule and in a typical radical pair in proteins. The radical electrons in proteins are spaced at a distance of about 1 nm, and the short-acting exchange interaction is small, since its magnitude is determined by the overlap of the wave functions of two electrons. Wave functions decrease exponentially with increasing distance between electrons, and for paramagnetic electrons in proteins, the exchange interaction is usually neglected. In an oxygen molecule, two electrons are localized near the same nucleus, so their wave functions overlap significantly. Here, the exchange interaction of electrons is large and leads — according to the Pauli prohibition principle — to a significant energy gap between C- and T-terms, on the order of 1 eV. As a result, magnetic effects with respect to the CT states of molecular oxygen begin to manifest in MP of the order 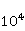 of E ( of E (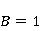 Tl) [78], when the energy of the magnetic moment of the electron is comparable to the gap value. Weak MP of the order of geoMP is 4-5 orders of magnitude smaller and does not have a direct effect on the oxygen molecule. Tl) [78], when the energy of the magnetic moment of the electron is comparable to the gap value. Weak MP of the order of geoMP is 4-5 orders of magnitude smaller and does not have a direct effect on the oxygen molecule. The mechanism of radical pairs In contrast to the effect of a strong MP on an oxygen molecule, the effect of MP on a pair of intermediate-state radicals takes place in much smaller fields, starting from several Mt. Therefore, the spin-chemical mechanism, RPM, is considered today the main candidate for explaining the effects of weak MP.
Its main advantage is that it certainly exists and successfully agrees with some of the patterns observed in magnetic navigation. These are the following patterns: 1) the dependence of the response to MP on the presence of optical radiation and its spectral composition, 2) invariance to the change of MP to the opposite direction, and 3) the absence of pronounced frequency selectivity in the low frequency range of variable MP. It is believed that radiation from the blue region of the optical spectrum leads to a photoinduced casting of an electron to one of the upper levels, followed by its transfer to another center and, thus, the birth of an electron pair of radicals. The quantum mechanical model of RPM predicts that the effect is a function of the square of the magnitude of MP [79] and, therefore, is invariant to the inversion of MP. The main disadvantage of RPM is its low sensitivity to MP. This still prevents us from unconditionally recognizing that the spin-chemical mechanism explains the response of organisms to MP at the geoMP level and, moreover, at the level of tens of nTl. It was noted above that the magnetic sense of some animals is combined with the visual apparatus. Let's take a closer look at how the RPM is embedded in the visual system. The visual photoreceptor is a "cone" or "wand" located in the retina of the eye and familiar from school, containing the protein cryptochrome. On the one hand, with the help of this protein, the light signal is converted into a biochemical signal. On the other hand, such a transformation involves the birth of pairs of radicals, and they are sensitive to MP. Over billions of years of evolution, photoreceptors of some birds have become involved in magnetoreception [32]. It is believed that photons of the blue spectral range initiate the birth of flavin-tryptophan radical pairs, whose spin and, consequently, chemical evolution depends on MP. Many details are not yet known, but the fact itself has been confirmed by numerous publications in highly rated journals, see e.g. [33,80,81]. An explanation of the effect of MP on photosynthetic bacteria was proposed in [82]. The population of the triplet state of a pair of photoinduced radicals in the reaction center depends on the magnitude of the MP. This affects photosynthesis. A year later, the authors [79] suggested that the orientation of birds in geoMP occurs using a quantum chemical compass. The CT conversion of radicals, taking into account the anisotropic hyperfine interaction, also depends on the direction (but not on the reverse) of the MP, which is consistent with the experiment [21]. Such mechanisms have received the collective name RPM. This mechanism, however, has a low sensitivity. In a separate photoreceptor, a change in MP by an order of magnitude of geoMP causes an effect that is unlikely to exceed 0.1% theoretically, and experimentally by orders of magnitude less [62,83]. The low sensitivity of the RPM is due to the short lifetime of the correlated state of the spins — 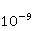 c, rarely c, rarely 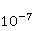 C. This is the time of thermal relaxation of the phase coherence of electron spins. In a short time, the MP should have time to noticeably change the state of a pair of spins relative to each other. The magnitude of this MP can be estimated by comparing the width of the quantum level associated with the time of thermal relaxation and the size of the Zeeman splitting of levels in MP. It is shown in [27,66] that qualitative changes in the dynamics of moments occur when the gap C. This is the time of thermal relaxation of the phase coherence of electron spins. In a short time, the MP should have time to noticeably change the state of a pair of spins relative to each other. The magnitude of this MP can be estimated by comparing the width of the quantum level associated with the time of thermal relaxation and the size of the Zeeman splitting of levels in MP. It is shown in [27,66] that qualitative changes in the dynamics of moments occur when the gap 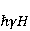 between the Zeeman sublevels becomes comparable to the width between the Zeeman sublevels becomes comparable to the width 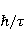 of the levels themselves. Consequently, the sensitivity of a biophysical sensor based on RPM can occur where there is a fundamental relationship of the levels themselves. Consequently, the sensitivity of a biophysical sensor based on RPM can occur where there is a fundamental relationship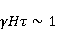 . The parameter . The parameter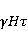 , among many others, controls the probability of the sensor's response to the MP. Thus, the MP, starting from which magnetic effects can become noticeable, is , among many others, controls the probability of the sensor's response to the MP. Thus, the MP, starting from which magnetic effects can become noticeable, is 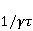 , where is the thermal relaxation time of the moment. For electrons with , where is the thermal relaxation time of the moment. For electrons with 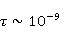 c, for example, in radicals of enzyme-substrate complexes, this value is 5 MT, which is a hundred times more than geoMP. For sensitivity at least at the level of 5 µT observed in non-specific effects, the relaxation time of the phase coherence of the spins of the radical pair should exceed 1 microsecond! It is not yet clear whether conditions could exist in living tissue at all to ensure such a weak connection of the electron with the thermostat [84,85]. Some authors believe [86] that the observation of a biological reaction in such weak MP proves that long spin relaxation times of electrons really exist. c, for example, in radicals of enzyme-substrate complexes, this value is 5 MT, which is a hundred times more than geoMP. For sensitivity at least at the level of 5 µT observed in non-specific effects, the relaxation time of the phase coherence of the spins of the radical pair should exceed 1 microsecond! It is not yet clear whether conditions could exist in living tissue at all to ensure such a weak connection of the electron with the thermostat [84,85]. Some authors believe [86] that the observation of a biological reaction in such weak MP proves that long spin relaxation times of electrons really exist. The authors [87,88] tried to justify large spin relaxation times theoretically. However, there are still no experiments confirming the existence of such a long relaxation time, with the exception of exotic fullerene-type systems that are not related to biology. The actual spin relaxation time in radical pairs can be obtained from measurements of the EPR line width in geoMP or by measurements of spin magnetic effects in vitro and their comparison with calculations. As far as we know, EPR signals in geoMP from the electron spins of intermediate radical pairs have not been observed in biochemical reactions. At the same time, the experience of spin chemistry, see e.g. [83], consistent with the theory, indicates a relatively fast spin relaxation under the conditions under discussion, about 1 ns. Perhaps 10 ns at most. Magnetic effects decrease approximately in proportion to the square of the spin relaxation time — if the rate of the chemical reaction is not high and does not suppress the magnetic effect to an even greater extent. Therefore, the question of the relaxation time of spin coherence is equivalent to the question of the existence of magnetic effects by the RPM mechanism in its standard form. Spin coherence is one of the mandatory conditions for the operation of quantum computers — a condition limiting their efficiency. Therefore, some authors study the relevance of describing RPM magnetoreception in terms typical of quantum computers - entanglement of quantum states and the distinction between pure and mixed states [89]. It is not yet clear whether such an expansion of the terminology base can give something new to solve the problems of magnetobiology. The concept of magnetic navigation based on spin-chemical effects in cryptochromes predicts effects that are qualitatively consistent with a significant part of observations and experiments. At the moment, this concept is a well-founded hypothesis, not devoid, however, of the above difficulty. Statistical amplification of weak magnetic signals
As follows from the material presented above, spin chemistry — or, in a more general context, the dynamics of magnetic moments — necessarily underlies the observed biological effects of weak MP. On the other hand, the analysis of the above quantum mechanisms shows that the magnitude of the primary physical changes, in particular in the RPM, is insufficient to reliably explain the results of experiments and observations. Apparently, small primary changes occurring as a result of the action of MP must be somehow "enhanced" in order to then cause tangible changes in the body at the level of concentrations of biochemical agents. This idea is not new. An enzymatic reaction highly sensitive to the concentration of the enzyme [90] and a chemical reaction in a mode close to bifurcation instability were proposed as an amplifying mechanism, when the reaction pathway can change abruptly with a slight variation in the concentration of the reactant [91-93]. These approaches have not been developed, probably due to the impossibility of their experimental verification in vivo. To explain the magnetic sense of some animals, the idea of statistical amplification of very small magnetic signals from millions of photoreceptors simultaneously entering the brain was attracted. The gain factor was insufficient to explain the magnetic navigation of animals based on RPM [94]. Moreover, this mechanism is not sufficient in relation to non—specific effects - in the absence of evolutionarily developed magnetoreceptors. However, as noted above, the very concept of statistical amplification, or accumulation, of small regular deviations against the background of fluctuations is attractive. This concept of detecting weak signals works wherever the concepts of a random variable, its average and variations are relevant — it is well-known and universal. It would be expected that magnetic effects, if they are caused by primary small deviations under the action of MP, could be detected where they accumulate in a long sequence of elementary acts. Let's pay attention to the fact that cyclic processes of biopolymer synthesis are ideal biochemical integrators, so that small changes in individual reaction cycles can accumulate and lead to the observed biological effects. The biological response to primary magnetic signals occurs due to integration in the biochemical system of the body. Of particular interest is the occurrence of errors in these processes. Even one mistake in any particular cycle can lead to the malfunction of the whole molecule of the synthesized biopolymer. We show how a small primary magnetic signal could be statistically enhanced along with the accumulation of the probability of biopolymerization errors [95]. The processes of synthesis of biopolymers are diverse. These are DNA replication, transcription — synthesis of complementary RNA, splicing, translation — synthesis of protein from amino acids in accordance with the mRNA code and post-translational folding of the protein chain into a globule and its maturation. Random errors may occur at each of these stages, but with significantly different probabilities. There are perfect biochemical mechanisms for correcting replication and transcription errors; their probability is therefore very small, of the order 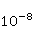 of and of and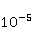 , respectively [96]. Translation errors occur much more often, with a probability of the order , respectively [96]. Translation errors occur much more often, with a probability of the order 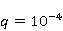 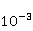 of one attached molecule of the translated amino acid [97,98]. About ten percent of synthesized molecules from several hundred links contain at least one error. For such molecules, the chance to adopt a native conformation in the folding process is significantly reduced. of one attached molecule of the translated amino acid [97,98]. About ten percent of synthesized molecules from several hundred links contain at least one error. For such molecules, the chance to adopt a native conformation in the folding process is significantly reduced. Errors also occur at the folding stage — one of the reasons is the intricate geometry and topological nodes of the folding trajectories [99]. It is believed, however, that misfolding is more likely to occur due to broadcast errors, rather than folding itself. Consequently, mainly errors in the translation of genetic information control the accuracy of gene expression. It is obvious that the possible influence of MP on the probability of translation errors is exactly the process where magnetic effects could manifest themselves at the biological level. This is also indirectly evidenced by the results of [100], where weak electromagnetic radiation affected gene expression without causing DNA breaks. Recently, such a mechanism of statistical amplification has been proposed, based on the fact that MP could affect not the concentration of native proteins necessary for the functioning of the body, but the concentration of non-functional proteins resulting from biopolymer synthesis errors [95]. Non-functional proteins can be cytotoxic [101] and in any case cause an additional load on the adaptive resources of the body.
Translation is a complex multi—stage cyclic process involving a variety of enzyme reactions. Translation is carried out by the ribosome, a special macromolecular machine for assembling amino acids into proteins. A simplification illustrating the occurrence of a noticeable error is as follows. Suppose there is a process of synthesis of a protein chain from a large number  of links with an equal probability of links with an equal probability 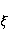 of inclusion error — the appearance of an incorrect amino acid residue in some place of the synthesized chain that does not correspond to the instructions of the mRNA. A native functional protein globule implies the absence of local errors in all parts of the chain. Then the probability of inclusion error — the appearance of an incorrect amino acid residue in some place of the synthesized chain that does not correspond to the instructions of the mRNA. A native functional protein globule implies the absence of local errors in all parts of the chain. Then the probability 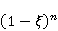 is the probability of the appearance of the "correct" amino acid sequence, and is the probability of the appearance of the "correct" amino acid sequence, and 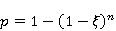 there is a probability of the appearance of a defective molecule, i.e. the probability of a translation error. The sensitivity of a quantity there is a probability of the appearance of a defective molecule, i.e. the probability of a translation error. The sensitivity of a quantity  to the probability of a local inclusion error is a derivative to the probability of a local inclusion error is a derivative 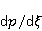 . Its magnitude is maximum under the condition . Its magnitude is maximum under the condition 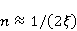 and can reach large values and can reach large values 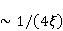 at small . The probability of the correct translation of the entire molecule in any case does not exceed one. This means that the effect of changing the probability of translation error when varying the probability of a local failure is that a change of several tenths occurs already with very small variations at small . The probability of the correct translation of the entire molecule in any case does not exceed one. This means that the effect of changing the probability of translation error when varying the probability of a local failure is that a change of several tenths occurs already with very small variations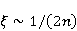 . . The number of different proteins in the human body exceeds two million [102], and their bulk has a length of one hundred to five hundred amino acids. Therefore, changes in the probability of local failures 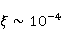 cause significant changes in the probability of the appearance of non–functional proteins. Note that levels cause significant changes in the probability of the appearance of non–functional proteins. Note that levels 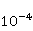 are the usual magnitude of relative effects in RPM. On the one hand, ribosomal translation is carried out with the help of special enzymes, and many enzymes work with a pronounced electron transfer stage. On the other hand, during electron transfer, an intermediate magnetosensitive state of a pair of radicals with spin-correlated electrons may occur. Indeed, the activity of some enzymes depends on MP, see e.g. [76,103]. Thus, the assumption of the involvement of RPM, or, in general, the dynamics of magnetic moments, in ribosomal translation is well-founded, and the mechanism of statistical amplification of translation errors consistently explains the nonspecific effect of weak MP on the body, Fig. 5. are the usual magnitude of relative effects in RPM. On the one hand, ribosomal translation is carried out with the help of special enzymes, and many enzymes work with a pronounced electron transfer stage. On the other hand, during electron transfer, an intermediate magnetosensitive state of a pair of radicals with spin-correlated electrons may occur. Indeed, the activity of some enzymes depends on MP, see e.g. [76,103]. Thus, the assumption of the involvement of RPM, or, in general, the dynamics of magnetic moments, in ribosomal translation is well-founded, and the mechanism of statistical amplification of translation errors consistently explains the nonspecific effect of weak MP on the body, Fig. 5. 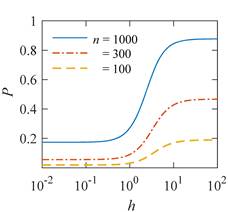
Fig. 5. An example of calculating the dependence of the average probability 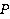 of ribosomal translation error on the relative value of MP of ribosomal translation error on the relative value of MP 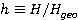 for different sizes for different sizes 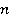 of the synthesized protein chain, according to [95]. of the synthesized protein chain, according to [95]. This mechanism of statistical amplification removes the main contradiction in the explanation of nonspecific magnetic effects based on RPM — a very small amount of RMP effects of the order of –. Such insignificant primary magnetic effects are amplified approximately by 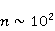 – –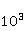 times and bring the total effect to the observed level. times and bring the total effect to the observed level. Hypomagnetic field as a research tool The study of biopolymer processes in a hypomagnetic field (hypoMP — MP, reduced in comparison with geoMP by two orders of magnitude or more) is a promising tool for identifying the molecular mechanism of the effect of low-intensity MP on organisms and the subsequent use of precision weak MP for selective control of gene expression. Indeed, the basis of such an influence is the primary effect of MP on magnetic moments of the atomic-molecular scale. On the one hand, today the primary target of MP in organisms or aquatic systems has not been established. Fig. 5, for example, demonstrates the expected dependences of the probability of an effect by the mechanism of RPM, i.e. when the primary targets are electrons. On the other hand, the targets could be the magnetic moments of protons or other magnetic nuclei, as well as the magnetic moments of molecular rotations of charged groups. These types of moments have different gyromagnetic ratios and, in the case of their precession in MP, precess at different angular velocities. Therefore, the best tool for magnetic influence is one that affects all magnetic moments regardless of their gyromagnetic factor. This is the hypoMP. In such a field, the precession of all magnetic moments slows down. Quantum mechanisms of hypoMP effects involving magnetic moments with different gyromagnetic ratios are considered in [27] and demonstrate rich dependence on MP features.
However, it is difficult to conduct experiments using hypoMP in normal laboratory conditions, since fluctuations in MP in laboratories often reach several MCT. Here there is an analogy with the use of proton NMR in a liquid, when the width of the resonance line is very narrow, is – Gs. In a geoMP with fluctuations, the random displacement of the line would significantly exceed the width of the line, which would make it difficult to observe the resonance. This difficulty is removed by increasing the MP by 6 orders of magnitude, that is, by using fields of the order of 1-10 T in NMR, when fluctuations of MP in several ?TL practically do not shift the lines — they do not cause noticeable relative changes. Another obvious way to reduce the influence of MP fluctuations is their active compensation or shielding, Fig. 6. It is against the background of hypoMP that it would be possible to observe the effect of additionally organized precision MP on molecular processes in organisms. Observation of such effects in strong fields is impossible, since in them the precession of all magnetic moments is high-frequency and does not correspond to the time scale of natural biopolymer processes. HypoMP could in practice be as interesting as it is strong. 
Fig. 6. One of the two identical blocks of the hypomagnetic exposure system implemented at IOF RAS compensates for external MP and reduces fluctuations of homogeneous ambient MP by several thousand times. The instrumental potential of a hypoMP in comparison with a strong MP can be represented by analogy with the use of strong and weak optical radiation. The natural "biological" reference point is the intensity of daylight. Powerful light in the form of, for example, laser radiation finds many obvious applications. Very weak radiations — single photons — allow us to study fundamental quantum processes and find application in the development of quantum computers and quantum cryptography. HypoMP is, apparently, an important area of biophysical research. The use of hypoMP fields in molecular biology opens up interesting prospects comparable in versatility to NMR. If the mechanism of statistical amplification of translation errors induced by MP presented above passes experimental testing and proves to be correct, then selective control of the rate of protein production without the use of chemicals will also become real — with the help of a specially organized precision MP against the background of hypoMP. This would have clear prospects of application, in addition to research, in medicine, pharmacology and agricultural biotechnologies. Conclusion Quantum nonspecific effects of weak MP have a general and universal character — they always occur in all organisms. These effects are invisible when small. When they are not small and could be noticeable, they, by virtue of their randomness, disguise themselves as the result of the action of uncontrolled random factors. This manifests itself in the form of non—reproducibility, - heterogeneity of the array of measurements or specifically increased random spread. Unlike magnetoreception — the action of weak MP on specialized receptors of some organisms — magnetic nonspecific effects do not receive the attention of researchers. The molecular physical mechanisms of non-specific effects remain largely unclear, although the understanding of their nature is gradually deepening. Knowledge of molecular mechanisms would significantly reduce the level of randomness of non-specific effects, i.e. increase their reproducibility, and find many useful applications for them. Among the promising ones are mechanisms based on the quantum dynamics of magnetic moments, primarily the magnetochemical mechanism, or RPM. The low sensitivity of RPM in the region of weak MP is compensated by the possible inclusion of spin-correlated radical pairs in the work of biopolymer enzymes, in particular ribosomal translation enzymes. This makes it possible to increase the magnetic sensitivity of the RPM by two or three orders of magnitude due to a statistical increase in the probability of local errors of cellular translation. Thus, the main problem of magnetobiology can be considered solved, since a consistent explanation of the biological effects of weak MP appears. The mechanisms of mixing the quantum levels of magnetic moments and the molecular gyroscope also have a certain explanatory potential in relation to the effects of MP. This paper also lists mechanisms that have been proposed at various times to explain the nonspecific biological effects of weak MP, but have proved to be untenable for one reason or another. These include induction and resonance mechanisms, as well as mechanisms based on the dynamics of magnetic nanoparticles. In general, all mechanisms that use classical, rather than quantum, physics to describe microscopic processes are untenable, for example, those that use the idea of the Lorentz force. It is shown that the study of the effect of hypoMP on cellular processes under conditions of compensation or screening of laboratory magnetic fluctuations has good prospects for a variety of practical applications. Applications 1. The mechanism of mixing quantum levels In addition to the specific effects of magnetoreception, as mentioned above, there are also non—specific effects - the epiphenomenon of the vital activity of organisms in geoMP. Among the frequent properties of non—specific responses are, for example, sensitivity to the reverse of the MP direction and pronounced frequency selectivity. RPM is not able to explain such properties. In addition, there are dozens of studies in which the observed patterns of magnetic response also do not fit into the RPM scenario for one reason or another, see e.g. [52,104]. In particular, the magnetic response is observed when excited by photons of the red spectral range, whose energy is insufficient for the generation of radical pairs [35], or in general in the dark. To explain such effects, apparently, some other mechanism is needed. And a separate MP sensor should respond to a change in the MP sign at least. The insensitivity of the RPM to the reverse of the MP is due to the fact that the MP changes the dynamics of a pair of moments relative to each other. Hence, a scenario makes sense where the MP would change the dynamics of a single moment relative to the selected direction given by the local molecular environment [105]. This is the mechanism of mixing quantum levels [27,62,66]. The scenario where the MP changes the dynamics of a single moment relative to a local selected direction has attractive properties. He is able to explain, so far qualitatively, the features of non-specific effects and gives simple quantitative ratios for verification [66]. The mechanism is extremely abstract and general, so there simply cannot be alternatives among single-particle quantum mechanisms. His conclusions do not depend on the nature of magnetic moments, and are experimentally verifiable. When applied to spin magnetic moments, the mechanism is quantum, since spin is a property of microparticles that obeys the laws of quantum physics. In the special case of a uniaxial MP that remains collinear to any axis, it is permissible to describe the mechanism using a vector spin model within the framework of classical physics, which represents significant mathematical convenience.
As mentioned above, the parameter controls the probability of the sensor's response to the MP. The experimentally found MP value 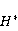 corresponding to the highest sensitivity is therefore associated with the product corresponding to the highest sensitivity is therefore associated with the product 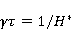 . And this allows us to clarify the nature of the primary target of MP. Indeed, there are few possible targets — an electron, a proton, another magnetic nucleus, the orbital moment of an electron or a charged molecular group — and . And this allows us to clarify the nature of the primary target of MP. Indeed, there are few possible targets — an electron, a proton, another magnetic nucleus, the orbital moment of an electron or a charged molecular group — and 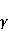 their values are significantly different and are often known by order of magnitude for different molecular environments. their values are significantly different and are often known by order of magnitude for different molecular environments. In addition, some of the primary sensors of the MP are located on rotating molecules [62]. Calculations show that the response of the rotating magnetic sensor to the MP is shifted relatively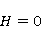 . In this case , the MP , now corresponding to the maximum response, is related to the speed of molecular rotation . In this case , the MP , now corresponding to the maximum response, is related to the speed of molecular rotation 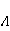 by the ratio by the ratio 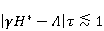 . It can be seen that the magnitude of the effect changes when the MP changes to the opposite direction, which is not present in the RMP, and which makes it possible to distinguish molecular mechanisms experimentally. Experimental determination of the dependence of the biological response on the MP value can provide information about the nature of the molecular processes of the nonspecific reaction of organisms to a weak MP. In particular, the authors of the experiments [35,106] interpret their results on the basis of the level mixing mechanism, considering the explanation based on the RPM unsatisfactory. . It can be seen that the magnitude of the effect changes when the MP changes to the opposite direction, which is not present in the RMP, and which makes it possible to distinguish molecular mechanisms experimentally. Experimental determination of the dependence of the biological response on the MP value can provide information about the nature of the molecular processes of the nonspecific reaction of organisms to a weak MP. In particular, the authors of the experiments [35,106] interpret their results on the basis of the level mixing mechanism, considering the explanation based on the RPM unsatisfactory. The description of the mechanism of mixing quantum levels requires, in contrast to the RPM, only a minimum number of variables and parameters. The disadvantage of the mechanism is the reverse side of its extreme abstractness, generality and universality. The mechanism says nothing about the nature of magnetic moments and the way they relate to the biophysical environment, although it allows you to gain new knowledge about the nature of moments from comparing your predictions with the experiment. 2. Molecular gyroscope The last scenario of the influence of MP on the magnetic moment is the influence on the magnetic moment of the spatial rotation of charged atomic—molecular groups, — on the orbital magnetic moment. The rotation of the charges leads to the appearance of a magnetic moment interacting with an external MP. MP changes the spatial structure or phases of angular quantum states of rotating molecular groups, which leads to interference effects [74] p. 385. Interference here means the heterogeneity of the probability density of different angular positions of the molecular group. The magnetic field, affecting the phases of quantum states, affects interference and, thereby, changes the probabilities of the presence of a molecular group in different angular positions. For example, a change in the probability of different angular positions of amino acid residues during the folding of a protein chain could affect the speed or results of folding. A rotating molecular group connected by two covalent bonds, or "supports", to the walls of the cavity formed by the molecular environment is a molecular gyroscope. Short segments of polypeptides or parts of nucleic acids could be considered as molecular gyroscopes sensitive to MP. For example, Watson-Crick pairs of nitrogenous bases that bind two strands of DNA into a double helix have a rotational degree of freedom. Their rotations are hindered due to steric limitations. However, in the field of DNA polymerase work, steric restrictions could be relaxed and allow for almost free rotation of pairs in a certain time interval. It is important that the thermal vibrations of the molecular gyroscope supports create only zero moments of forces relative to its own axis of rotation. This means that the gyroscopic degree of freedom relaxes slowly — it is immune to thermal vibrations of the supports. Estimates of thermalization due to van der Waals interactions carried out in [70] showed that if a realistic molecular gyroscope formed by an amino acid residue of phenylalanine is embedded in a cavity with a radius of 1.4 nm, then the relaxation time of its coherent rotation is 10 ms in order of magnitude. The relaxation time increases exponentially with the size of the cavity, and at a radius of 1.6 nm reaches 1 s. The gyromagnetic ratio of the molecular gyroscope is 5-6 orders of magnitude smaller than the electronic one due to the large mass of the gyroscope, but the coherence relaxation time, as can be seen, is 6-9 orders of magnitude longer than the electronic one. Therefore, the fundamental relation governing the occurrence of magnetic effects could be fulfilled here. It is not yet clear whether there are really molecular structures like a gyroscope that have a long relaxation time. On the one hand, the presence of the necessary and more or less water-free cavities with a radius of about 1.5 nm is problematic. On the other hand, such cavities do not necessarily have to exist as immutable structures. It is enough that they appear as virtual biophysical formations with a lifetime of 0.01–1 s. It is possible that such cavities could be formed during folding and maturation of long protein molecules, during various conformational transformations of chromatin-DNA complexes. The predictive power of the gyroscopic mechanism is the same as that of the quantum level mixing mechanism. Theoretical estimates show a rich behavior of the molecular gyroscope in variable MP, in constant MP and, in particular, in hypoMP conditions [27]. The disadvantage of the mechanism is that today it remains a speculative concept that has not been confirmed experimentally in any way. 3. Untenable mechanisms of biological effects of weak MP The following are magnetic phenomena that cannot underlie the nonspecific sensitivity of organisms to MP. Magnetic induction
This is the induction of eddy currents by high-frequency or pulsed MP. Typically, the eddy currents of pulsed MP significantly exceed the currents generated by MP in the magnetobiology of nonspecific effects. One of the ICNIRP standards [20] for the population sets the value of 2 mA/m as the main limitation of the current density as the main limitation of the current density  with a frequency of 4-1000 Hz in the human body. It can be assumed that noticeable effects occur where a current of slightly higher density passes through the body. Let's take with a frequency of 4-1000 Hz in the human body. It can be assumed that noticeable effects occur where a current of slightly higher density passes through the body. Let's take 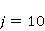 mA/m for estimates. For example, in [107], the transcription level in HL-60 cells varied up to 30% at a current density from 3 to 30 mA/m. Using Ohm's law mA/m for estimates. For example, in [107], the transcription level in HL-60 cells varied up to 30% at a current density from 3 to 30 mA/m. Using Ohm's law 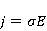 and Faraday's law of electromagnetic induction and Faraday's law of electromagnetic induction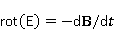 , where , where 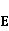 and and 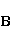 are vectors of electric field strength and magnetic flux density, it is easy to estimate the rate of change of MP in a centimeter sample with a conductivity are vectors of electric field strength and magnetic flux density, it is easy to estimate the rate of change of MP in a centimeter sample with a conductivity 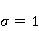 of Cm/m, at which current induction with a density of mA / m occurs. The speed is 1 T/s. In order for a sinusoidal 50 Hz MP to cause such rapid changes, it must have an amplitude of about 20 Mt. However, many magnetobiological effects occur in MP a thousand times smaller and even in constant MP that do not cause eddy currents. In these cases, eddy currents in the body, as well as heating, are insignificant or impossible. Thus, mechanisms based on the phenomenon of electromagnetic induction are not able to explain the molecular biological effects of weak low-frequency and, moreover, constant MP. of Cm/m, at which current induction with a density of mA / m occurs. The speed is 1 T/s. In order for a sinusoidal 50 Hz MP to cause such rapid changes, it must have an amplitude of about 20 Mt. However, many magnetobiological effects occur in MP a thousand times smaller and even in constant MP that do not cause eddy currents. In these cases, eddy currents in the body, as well as heating, are insignificant or impossible. Thus, mechanisms based on the phenomenon of electromagnetic induction are not able to explain the molecular biological effects of weak low-frequency and, moreover, constant MP. Electromagnetic induction should be taken into account when interpreting effects in high-frequency or pulsed MP. Since the probability of induction effects is proportional to the frequency of MP, the observed biological response may be mistakenly attributed to the magnetic effect. Magnetic resonance Magnetic resonance, both electronic and nuclear, cannot be an explanation for the nonspecific effects of MP. Magnetic resonance underlies, for example, medical tomography and scientific magnetic spectrometry. To observe magnetic resonance, special conditions are required: a strong static field of several Tl and a variable MP perpendicular to it with a high-frequency component determined by a static MP. The conditions of magnetobiological effects are far from these strict requirements. Other resonant phenomena are also excluded, where we are talking about the accumulation of weak MP energy in the vibrations of atoms and molecules. In a low-intensity MP, such accumulation, even under idealized conditions, would require coherent oscillations for months or more [22] p. 366, which is absurd. There is an assumption in the literature that fluctuations of the Earth's MP at the frequencies of Schumann resonances can cause a biological response in the human body due to coincidence with the frequencies 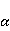 of other rhythms of brain activity. The surface of the Earth and the ionized layer of the atmosphere form a waveguide in which electromagnetic waves propagate. The frequency of natural oscillations of the field in such a waveguide is determined by the condition of multiplicity of the length of the electromagnetic wave and the circumference of the Earth. Resonances are excited by thunderstorm activity with intense lightning. V. Shuman calculated the resonance frequencies taking into account dissipation and spherical symmetry: 8, 14, 20 Hz, etc. The resonance is most pronounced at the lowest frequency of about 8 Hz, corresponding to the rhythm. of other rhythms of brain activity. The surface of the Earth and the ionized layer of the atmosphere form a waveguide in which electromagnetic waves propagate. The frequency of natural oscillations of the field in such a waveguide is determined by the condition of multiplicity of the length of the electromagnetic wave and the circumference of the Earth. Resonances are excited by thunderstorm activity with intense lightning. V. Shuman calculated the resonance frequencies taking into account dissipation and spherical symmetry: 8, 14, 20 Hz, etc. The resonance is most pronounced at the lowest frequency of about 8 Hz, corresponding to the rhythm. Resonant oscillations of the electromagnetic field can be detected on the Earth's surface by measurements of the electric or magnetic components. However, these vibrations cannot cause a magnetic biological response, since the coincidence of the resonance and rhythm frequencies alone is not enough. It is necessary, at least, that the magnitude of the field fluctuations be large enough. The magnitude of the Schumann oscillations of the MP on the Earth's surface is negligible, on the order of 1 pt. The MP will change by this value if, for example, you turn on a pocket flashlight ten meters from the place of measurement. This is several orders of magnitude less than the intensity of urban magnetic fluctuations of the same frequencies [22] p. 202. The uselessness of this concept is also evident in the fact that today the nature of the biological action of MP of the order of 1 µT, i.e. fields a million times stronger than Schumann magnetic signals, is not clear. Magnetic nanoparticles The possible involvement of magnetic nanoparticles in non-specific magnetic effects is often discussed. Indeed, biogenic magnetite particles up to tens of nanometers in size have been found in many organisms. Their energy in GEOMPS often exceeds the characteristic scale of the activation energy of chemical reactions [108,109], where the Boltzmann constant is the absolute temperature. In MP, nanoparticles are subjected to torque and could compress nearby biophysical structures. However, with respect to non-specific effects, this idea has not yet found either theoretical [27] or experimental confirmation. In addition, the magnetic effect is also observed in cultures where magnetic nanoparticles are deliberately excluded — in various bacteria, in animal and plant cell cultures. For this reason, mechanisms based on the dynamics of magnetic nanoparticles cannot underlie non-specific magnetic effects. The Lorentz Force There is a certain pattern in the chaotic movement of free ions in biological tissues in MP. Under the influence of the Lorentz force — perpendicular to the velocity vector and the MP vector — the movement of the ion in the intervals between collisions with the molecules of the medium ceases to be rectilinear. The movement is carried out in small arcs, so that on average there is a general circular motion of ions. Such a movement creates a very weak MP directed opposite to the applied one. In other words, the diamagnetic moment of the medium increases. However, this effect is extremely small [22] p. 236, orders of magnitude less than electron diamagnetism. Therefore, the Lorentz force is impossible as a cause of magnetic biological effects in a weak MP.
References
1. Middendorff, A. T. Die Isepiptesen Russlands. (1855). Kaiserlichen Akademie der Wissenschaften.
2. Schott, H. Zur. (1996). Geschichte der Elektrotherapie und ihrer Beziehung zum Heilmagnetismus. In Naturheilverfahren und Unkonventionelle Medizinische Richtunge (Ed. V. Fialka und andere). Springer.
3. Kholodov, Yu. A. (1970). Magnetism in Biology. Nauka.
4. Kholodov, Yu. A. (1991). Bypassing the Sensory Organs? Znanie.
5. Presman, A. S. (1965). The action of microwaves on living organisms and biological structures. Physics-Uspekhi 8, 463–488.
6. Webb, S. J., Dodds, D. E. (1968). Microwave inhibition of bacterial growth. Nature 218, 374–375.
7. Devyatkov, N. D. (1974). Influence of millimeter-band electromagnetic radiation on biological objects. Physics–Uspekhi 16, 568–569.
8. Tambiyev, A. H., & Kirikova, & N. N., & Betskii, & O. V., Gulyaev, Y. V. (2003). Millimeter Waves and Photosynthetic Organisms. Radioengineering.
9. Belpomme, D., & Hardell, L., & Belyaev, & I., & Burgio, E., & Carpenter, D. O. (2018). Thermal and non-thermal health effects of low intensity non-ionizing radiation: An international perspective. Environmental Pollution 242, 643–658.
10. Chuyan, E. N., & Djeldubaeva, E. R. (2020). Low-Intensity Millimeter-Wave Radiation: Neuro Immunoendocrine Mechanisms of Adaptative Reactions. IT ARIAL.
11. Presman, A. S. (1968). Electromagnetic Fields and Wildlife. Nauka.
12. Anikin, V. M. (2016). Magnetobiology and magnetotherapy: Karmilov period. Heteromagnetic Microelectronics 21, 131–140.
13. Moulder, J. E. (2000). The electric and magnetic fields research and public information dissemination (EMF-RAPID) program. Radiation Research 153, 613–616.
14. IARC. (2002). IARC monographs on the evaluation of carcinogenic risks to humans. Vol. 80. Non-ionizing radiation, part 1: Static and extremely low-frequency (ELF) electric and magnetic fields. IARC Press.
15. Huss, A., & Peters, S., & Vermeulen, R. (2018). Occupational exposure to extremely low-frequency magnetic fields and the risk of ALS: A systematic review and meta-analysis. Bioelectromagnetics 39, 156–163.
16. Miller, A. B., & Morgan, L. L., & Udasin, I., & Davis, D. L. (2018). Cancer epidemiology update, following the 2011 IARC evaluation of radiofrequency electromagnetic fields (Monograph 102). Environmental Research, 167, 673–683.
17. Li, D.-K., & Chen, H., & Ferber, J. R., & Odouli, R., & Quesenberry, Ch. (2017). Exposure to magnetic field non-ionizing radiation and the risk of miscarriage: A prospective cohort study. Scientific Reports 7, 17541.
18. Belyaev, I., & Dean, A., & Eger, H., & Hubmann, & G., Jandrisovits, R., & Kern, M., et al. (2016). EUROPAEM EMF Guideline 2016 for the prevention, diagnosis and treatment of EMF-related health problems and illnesses. Reviews on Environmental Health, 31, 363–397.
19. Environmental health criteria 238. (2007). Extremely low frequency fields. World Health Organization.
20. International Commission on Non-Ionizing Radiation Protection. (1998). Guidelines for limiting exposure to time-varying, electric, magnetic and electromagnetic fields up to 300 GHz. Health Physics 74, 494–521.
21. Wiltschko, W., & Wiltschko, R. (1972). Magnetic compass of european robins. Science, 176, 62–64.
22. Binhi, V. N. (2011). Principles of Electromagnetic Biophysics. Fizmatlit.
23. Roosli, M. (Ed.). (2014). Epidemiology of Electromagnetic Fields. CRC Press.
24. Kudryashov, Yu. B., & Rubin, A. B. (2014). Radiation Biophysics: Ultra-Low Frequency Electromagnetic Radiation. Fizmatlit.
25. Markov, M. S. (Ed.). (2015). Electromagnetic Fields in Biology and Medicine. CRC Press.
26. Greenebaum, B., & Barnes, F. (2019) Biological and Medical Aspects of Electromagnetic Fields. Vols. 1, 2. CRC Press.
27. Binhi, V. N., & Prato, F. S. (2017). Biological effects of the hypomagnetic field: An analytical review of experiments and theories. PLoS ONE, 12, e0179340.
28. Binhi, V. N. (2019). Nonspecific magnetic biological effects: A model assuming the spin-orbit coupling. The Journal of Chemical Physics, 151, 204101.
29. Lohmann, K. J., & Lohmann, C. M., & Ehrhart, L. M., & Bagley, D. A., & Swing, T. (2004). Animal behaviour: Geomagnetic map used in sea-turtle navigation. Nature, 428, 909–910.
30. Bianco, G., Ilieva, M., Åkesson, S. (2019). Magnetic storms disrupt nocturnal migratory activity in songbirds. Biology Letters, 15, 20180918.
31. Kishkinev, D. A., & Chernetsov, N. S. (2014). Magnetoreception systems in birds: A review of current research. General Biology Journal, 75, 104–123.
32. Hore, P. J., & Mouritsen, H. The radical-pair mechanism of magnetoreception. Annual Review of Biophysics, 45, 299–344 (2016).
33. Wan, G., & Hayden, A. N., Iiams, & S. E., Merlin, C. (2021). Cryptochrome 1 mediates light-dependent inclination magnetosensing in monarch butterflies. Nature Communications, 12, 771.
34. Novitsky, Yu. I., & Novitskaya, G. V. (2016). The Effect of a Constant Magnetic Field on Plants. Nauka.
35. Dhiman, S. K., & Wu, F., & Galland, P. (2022). Effects of weak static magnetic fields on the development of seedlings of arabidopsis thaliana. Protoplasma Sep 21.
36. Breus, T. K., & Binhi, V. N., & Petrukovich, A. A. (2016). Magnetic factor of the solar terrestrial relations and its impact on the human body: physical problems and prospects for research. Physics–Uspekhi, 59, 502–510.
37. Fu, J.-P., & Mo, W.-C., & Liu, Y., & He, R.-Q. (2016). Decline of cell viability and mitochondrial activity in mouse skeletal muscle cell in a hypomagnetic field. Bioelectromagnetics, 37, 212–222.
38. Paponov, I. A., & Fliegmann, J., & Narayana, R., & E., M. M. (2021). Differential root and shoot magnetoresponses in arabidopsis thaliana. Scientific Reports, 11, 1–15.
39. Johnsen, S., & Lohmann, K. J. (2005). The physics and neurobiology of magnetoreception. Nature Reviews. Neuroscience, 6, 703–712.
40. Buchachenko, A. L. (2014). Magnetic field-dependent molecular and chemical processes in biochemistry, genetics and medicine. Russian Chemical Reviews, 83, 1–12.
41. Yang, X., & Li, Zh., Polyakova, T., & Dejneka, A., & Zablotskii, V., & Zhang, X. (2020). Effect of static magnetic field on DNA synthesis: The interplay between DNA chirality and magnetic field left–right asymmetry. FASEB BioAdvances, 2, 254–263.
42. Schrödinger, E. (2002). What is Life? The Physical Aspect of the Living Cell. R&C Dynamics.
43. Binhi, V. N. (2021). Physical Effects of Consciousness: The Law of Reproducibility. Infra-M, www.lawr.info.
44. Ivanitskii, G. R. (2010). 21st century: What is life from the perspective of physics? Physics-Uspekhi, 53, 327–356.
45. Marais, A., & Adams, B., & Ringsmuth, A. K., & Ferretti, M., & Gruber, J. M., & Hendrikx, R., et al. (2018). The future of quantum biology. Journal of The Royal Society Interface, 15, 20180640.
46. Syurakshin, A. V., & Saleev, V. A., & Yushankhai, V. Yu. (2022). Quantum models in biology. Vestnik of Samara University. Natural Science Series, 28, 74–94.
47. Binhi, V. N., & Rubin, A. B. (2022). Theoretical concepts in magnetobiology after 40 years of research. Cells, 11, 274.
48. Sarimov, R. M., Binhi, V. N. (2020). Low-frequency magnetic fields in cars and office premises and the geomagnetic field variations. Bioelectromagnetics, 41, 360–368.
49. Makinistian, L., & Muehsam, D. J., & Bersani, F., & Belyaev, I. (2018). Some recommendations for experimental work in magnetobiology, revisited. Bioelectromagnetics, 39, 556–564.
50. Prato, F. S., & Desjardins-Holmes, D., & Keenliside, L. D., & DeMoor, J. M., & Robertson, J. A., & Thomas, A. W. (2013). Magnetoreception in laboratory mice: Sensitivity to extremely low frequency fields exceeds 33 nT at 30 Hz. Journal of The Royal Society Interface, 10.
51. Engels, S., & Schneider, N.-L., & Lefeldt, N., & Hein, C. M., & Zapka, M., & Michalik, A., et al. (2014). Anthropogenic electromagnetic noise disrupts magnetic compass orientation in a migratory bird. Nature, 509, 353–356.
52. Bojarinova, J., & Kavokin, K., & Pakhomov, A., & Cherbunin, R., & Anashina, A., & Erokhina, M., et al. (2020). Magnetic compass of garden warblers is not affected by oscillating magnetic fields applied to their eyes. Scientific Reports, 10, 3473.
53. Pishchalnikov, Y., R., & Gurfinkel, Yu. I., & Sarimov, R. M., & Vasin, A. L., & Sasonko, M. L., & Matveeva, T. A., et al. (2019). Cardiovascular response as a marker of environmental stress caused by variations in geomagnetic field and local weather. Biomedical Signal Processing and Control, 51, 401–410.
54. Krylov, V. V., & Kantserova, N. P., & Lysenko, L. A., & Osipova, E. A. (2019). A simulated geomagnetic storm unsynchronizes with diurnal geomagnetic variation affecting calpain activity in roach and great pond snail. International Journal of Biometeorology, 63, 241–246.
55. Janashia, K., & Tvildiani, L., & Tsibadze, T., & Invia, N. (2022). Effects of the geomagnetic field time-varying components compensation as evidenced by heart rate variability of healthy males. Life Sciences in Space Research, 32, 38–44.
56. Binhi, V. N. (2021). Random effects in magnetobiology and a way to summarize them. Bioelectromagnetics, 42, 501–515.
57. Zenchenko, T. A., & Breus, T. K. (2023). Possible causes for the instability of reproduction of heliobiological results. Physics of Biology and Medicine.
58. Valberg, P. A. (1995). Designing EMF experiments: What is required to characterize ‘exposure’? Bioelectromagnetics, 16, 396–401.
59. Buchachenko, A. (2016). Why magnetic and electromagnetic effects in biology are irreproducible and contradictory? Bioelectromagnetics, 37, 1–13.
60. Portelli, L. (2019). Overcoming the irreproducibility barrier. in Bioengineering and biophysical aspects of electromagnetic fields (eds. Greenebaum, B., Barnes, F.) pp. 435–462. CRC Press.
61. Koonin, E. V. (2012). The Logic of Chance: The Nature and Origin of Biological Evolution. FT Press.
62. Binhi, V. N., & Prato, F. S. (2018). Rotations of macromolecules affect nonspecific biological responses to magnetic fields. Scientific Reports, 8, 13495.
63. Zenchenko, T. A., & Breus, T. K. (2021). The possible effect of space weather factors on various physiological systems of the human organism. Atmosphere, 12, 1–28.
64. Krylov, V. V., & Bolotovskaya, & I. V., Osipova, & E. A. (2013). The response of European Daphnia magna Straus and Australian Daphnia carinata King to changes in geomagnetic field. Electromagnetic Biology and Medicine, 32, 30–39.
65. Vasin, A. L., & Shafirkin, A. V., & I., G. Yu. (2019). Effect of artificial alternating geomagnetic field in the millihertz range on the heart rate variability indices. Aerospace and Environmental Medicine, 53, 62–69.
66. Binhi, V. N., & Prato, F. S. (2017). A physical mechanism of magnetoreception: extension and analysis. Bioelectromagnetics, 38, 41–52.
67. Buchachenko, A. (2014). Magneto-Biology and Medicine. Nova Science.
68. Binhi, V. N. 1995. Nuclear spins in the primary mechanisms of biological action of magnetic fields. Biophysics, 40, 671–685.
69. Binhi, V. N. (1998). On structural defects of liquid water in magnetic and electric fields. Biomedical Radioelektronics, 7, 16.
70. Binhi, V. N., & Savin, A. V. (2002). Molecular gyroscopes and biological effects of weak extremely low-frequency magnetic fields. Physical Review, E 65, 051912.
71. Fesenko, E. E., & Gluvstein, A. Ya. (1995). Changes in the state of water, induced by radiofrequency electromagnetic fields. FEBS Lett., 367, 53–55.
72. Lobyshev, V. I. (2005). Water is a sensor to weak forces including electromagnetic fields of low intensity. Electromagnetic Biology and Medicine, 24, 449–461.
73. Binhi, V. N., & Rubin, A. B. (2007). Fundamental problem of magnetobiology. Biomedical Technologies and Radioelectronics, 63, 76.
74. Binhi, V. N. (2002). Magnetobiology: Underlying Physical Problems. Academic Press.
75. Afanasyeva, M. S., & Taraban, M. B., & Purtov, P. A., & Leshina, T. V., & Grissom, C. B. (2006). Magnetic spin effects in enzymatic reactions: Radical oxidation of NADH by horseradish peroxidase. Journal of the American Chemical Society, 128, 8651–8658.
76. Buchachenko, A. L., & Kuznetsov, D. A., & Breslavskaya, N. N. (2012). Chemistry of enzymatic ATP synthesis: An insight through the isotope window. Chemical Reviews, 112, 2042–2058.
77. Minaev, B. F. (2007). Electronic mechanisms of molecular oxygen activation. Russian Chemical Reviews, 76, 989–1011.
78. Amonkosolpan, J., & Aliev, G. N., & Wolverson, D., & Snow, P. A., & Davies, J. J. (2014). Magnetic field dependence of singlet oxygen generation by nanoporous silicon. Nanoscale Research Letters, 9, 342.
79. Schulten, K., & Swenberg, C., & Weller, A. (1978). A biomagnetic sensory mechanism based on magnetic field modulated coherent electron spin motion. Zeitschrift Fur Physikalische Chemie, 111, 1–5.
80. Xu, J., & Jarocha, L. E., & Zollitsch, T., & Konowalczyk, M., & Henbest, K. B., & Richert, S., et al. (2021). Magnetic sensitivity of cryptochrome 4 from a migratory songbird. Nature, 594, 535–540.
81. Mouritsen, H. (2018). Long-distance navigation and magnetoreception in migratory animals. Nature, 558, 50–59.
82. Hoff, A. J., & Rademaker, H., & Van Grondelle, & R., Duysens, L. N. M. (1977). On the magnetic field dependence of the yield of the triplet state in reaction centers of photosynthetic bacteria. Biochimica Et Biophysica Acta – Bioenergetics, 460, 547–554.
83. Steiner, U. E., & Ulrich, T. (1989). Magnetic field effects in chemical kinetics and related phenomena. Chemical Reviews, 89, 51–147.
84. Fedin, M. V., & Shakirov, S. R., & Purtov, P. A., & Bagryanskaya, E. G. (2006). Electron spin relaxation of radicals in weak magnetic fields. Russian Chemical Bulletin, 55, 1703–1716.
85. Ivanov, K. L., & Petrova, M. V., & Lukzen, N. N., & Maeda, K. (2010). Consistent treatment of spin-selective recombination of a radical pair confirms the Haberkorn approach. Journal of Physical Chemistry A, 114, 9447–9455.
86. Gauger, E. M., & Rieper, E., & Morton, J. J. L., & Benjamin, S. C., & Vedral, V. (2011). Sustained quantum coherence and entanglement in the avian compass. Physical Review Letters, 106, 040503.
87. Kattnig, D. R., & Solov’yov, I., & Hore, P. J. (2016). Electron spin relaxation in cryptochrome-based magnetoreception. Physical Chemistry Chemical Physics, 18, 12443–12456.
88. Worster, S., & Kattnig, D. R., & Hore, P. J. (2016). Spin relaxation of radicals in cryptochrome and its role in avian magnetoreception. The Journal of Chemical Physics, 145, 035104.
89. Cai, J., & Plenio, M. B. (2013). Chemical compass model for avian magnetoreception as a quantum coherent device. Physical Review Letters, 111, 230503.
90. Weaver, J. C., & Vaughan, T. E., & Astumian, R. D. (2000). Biological sensing of small field differences by magnetically sensitive chemical reactions. Nature, 405, 707–709.
91. Grundler, W., & Kaiser, F., & Keilmann, F., & Walleczek, J. (1992). Mechanisms of electromagnetic interaction with cellular systems. Naturwissenschaften, 79, 551–559.
92. Riznichenko, G. Yu., & Plyusnina, T. Yu., & Aksyonov, S. I. (1994). Modelling of the effect of a weak electric field on a nonlinear transmembrane ion transfer system. Bioelectrochem. Bioenerg., 35, 39–47.
93. Player, T. C., & Baxter, E. D. A., & Allatt, S., & Hore, P. J. (2021). Amplification of weak magnetic field effects on oscillating reactions. Scientific Reports, 11, 9615.
94. Binhi, V. N. (2023). Magnetic navigation of animals, contrast sensitivity of vision, and the Weber-Fechner law. Sensory Systems, 37, 46–60.
95. Binhi, V. N. (2023). Statistical amplification of the effects of weak MFs in cellular translation. Cells, 12, 724.
96. Mohler, K., & Ibba, M. (2017). Translational fidelity and mistranslation in the cellular response to stress. Nature Microbiology, 2, 17117.
97. Parker, J. (1989). Errors and alternatives in reading the universal genetic code. Microbiological Reviews, 53, 273–298.
98. Kurland, C. G. (1992). Translational accuracy and the fitness of bacteria. Annual Review of Genetics, 26, 29–50.
99. Nissley, D. A., & Jiang, Y., Trovato, F., Sitarik, I., Narayan, K. B., To, P., et al. (2022). Universal protein misfolding intermediates can bypass the proteostasis network and remain soluble and less functional. Nature Communications, 13, 3081.
100. Belyaev, I. Y., & Koch, C. B., & Terenius, O., & Roxstrom-Lindquist, & K., Malmgren, & L. O. G., Sommer, W. H., et al. (2006). Exposure of rat brain to 915 MHz GSM microwaves induces changes in gene expression but not double stranded DNA breaks or effects on chromatin conformation. Bioelectromagnetics, 27, 295–306.
101. Drummond, D. A., & Wilke, C. O. (2008). Mistranslation-induced protein misfolding as a dominant constraint on coding-sequence evolution. Cell, 134, 341–352.
102. Legrain, P., & Aebersold, R., & Archakov, A., & Bairoch, A., & Bala, K., & Beretta, L., et al. (2011). The human proteome project: Current state and future direction. Molecular & Cellular Proteomics, 10.
103. Stovbun, S. V., & Zlenko, D. V., & Bukhvostov, A. A., & Vedenkin, A. A., & Skoblin, A. A., & Kuznetsov, D. A., et al. (2023). Magnetic field and nuclear spin influence on the DNA synthesis rate. Scientific Reports, 13, 465.
104. Agliassa, C., & Maffei, M. E. (2019). Reduction of geomagnetic field (GMF) to near null magnetic field (NNMF) affects some Arabidopsis thaliana clock genes amplitude in a light independent manner. Journal of Plant Physiology, 232, 23–26.
105. Binhi, V. N. (2016). Primary physical mechanism of the biological effects of weak magnetic fields. Biophysics, 61, 170–176.
106. Dhiman, S. K., & Galland, P. (2018). Effects of weak static magnetic fields on the gene expression of seedlings of Arabidopsis thaliana. Journal of Plant Physiology, 231, 9–18.
107. Blank, M., & Soo, L., & Lin, H., Henderson, A. S., & Goodman, R. (1992). Changes in transcription in HL-60 cells following exposure to alternating currents from electric fields. Bioelectrochemistry and Bioenergetics, 28, 301–309.
108. Binhi, V. N., & Chernavsky, D. S. (2005). Stochastic resonance of magnetosomes fixed in the cytoskeleton. Biophysics, 50, 599–603.
109. Kirschvink, J. L., & Winklhofer, M., & Walker, M. M. (2010). Biophysics of magnetic orientation: Strengthening the interface between theory and experimental design. Journal of the Royal Society Interface 7, 179–171.
Link to this article
You can simply select and copy link from below text field.
|
|



